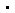2024年9月23日,北京大学王继纵课题组与邓兴旺课题组合作在cell发表了题为“light-induced remodeling of phytochrome b enables signal transduction by phytochrome-interacting factor”的研究论文,通过单颗粒冷冻电镜结构解析、phyb突变体蛋白光谱测定、结合生化实验表征、以及转基因植物表型分析,对模式植物拟南芥光激活态光敏色素phyb结构和功能进行了深入分析,阐明了其光激活变构机理,并在此基础上提出了phyb光信号转导的最初反应机制。
高等植物主要编码以phya和phyb为代表的两类光敏色素。phya属于i型光敏色素(光敏感型),主要在幼苗见光之前的黄化苗阶段大量存在,见光之后迅速被降解,主要作为远红光受体发挥功能;而phyb则属于ii型光敏色素(光不敏感型),在光下也可以稳定存在,是介导可逆红光响应的主要红光受体。phya或phyb都是由脱辅基蛋白二聚体共价结合四吡咯发色团(phytochromobilin, pφb)的形式存在。光敏色素通过发色团pφb在红光吸收态(pr,基态)以及远红光吸收态(pfr,激活态)之间进行可逆转变。拟南芥phyb被光激活后,可以直接与一类光敏色素互作因子(phytochrome-interacting factor,pif)互作,传递光信号并调控下游基因表达,促进光形态建成。因此phyb和pifs构成了植物响应周围光环境的关键信号模块。拟南芥中8个pif成员(pif1-8)均包含两个重要结构域:n端为结合phyb-pfr的激活态结合域(active-phyb binding motif,apb),c端为结合dna的bhlh二聚化结构域。在黑暗条件下,phyb-pr定位于细胞质中,pif1/3/4/5在核中作用驱动幼苗暗形态建成的发育过程(下胚轴伸长,子叶闭合)。一旦幼苗感知红光,phyb-pfr入核进而负调控上述pifs,抑制下胚轴伸长和促进子叶展开,并维持植物光生长形态。尽管2022年美国vierstra团队在nature发文揭示了phyb-pr的结构,但phyb-pfr及其识别pif的结构生物学基础及其调控机理仍然未知。

图1. 植物光敏色素的紫外-可见光吸收谱图(引自li, et al. 2011)
这项研究首次报道了光激活态光敏色素phyb-pfr以及不依赖于光的组成型激活突变体phyby276h分别结合下游信号分子pif6的复合物高分辨率冷冻电镜结构。首次发现光激活导致phyb-pr发生大规模结构重排,即phyb感光模块(photosensory module, psm)从pr状态下“头对尾”二聚体转变为pfr状态下“头对头”二聚体,且只能结合一个pif6-apb单体,最终形成phyb-pif6不对称三聚体(图2)。
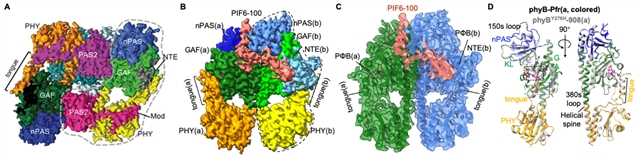
图2. phyb-pfr-pif6(b)和phyby276h-pif6(c)均为不对称三聚体,整体结构几乎完全一致(d),且均与文献报道的phyb-pr(a)二聚化方式存在明显差异
基于结构发现和实验表征,作者揭示了phyb光激活变构的详细分子机制。红光照射导致发色团小分子pφb构象变化,进一步引起phy结构域舌状突出结构由β片层向α螺旋的构象转变,新形成的α螺旋破坏了phyb-pr“头对尾”二聚体的稳定性,导致phyb构象的进一步重塑,最终形成的phyb-pfr结构提供了结合pif-apb的可能。随即一分子pif通过“诱导契合”的方式结合至phyb-nte,同时进一步维持了phyb-pfr“头对头”二聚体的稳定,最终形成phyb-pif信号复合物(图3)。
此外,作者报道的phyby276h-pif6复合物结构中,pφb虽然不具备光激发变构驱动phyb蛋白构象变化的能力,但它对于phyby276h始终保持pfr类似构象必不可少。结构比对表明phyby276h-pif6复合物结构与phyb-pfr-pif6几乎完全一致。
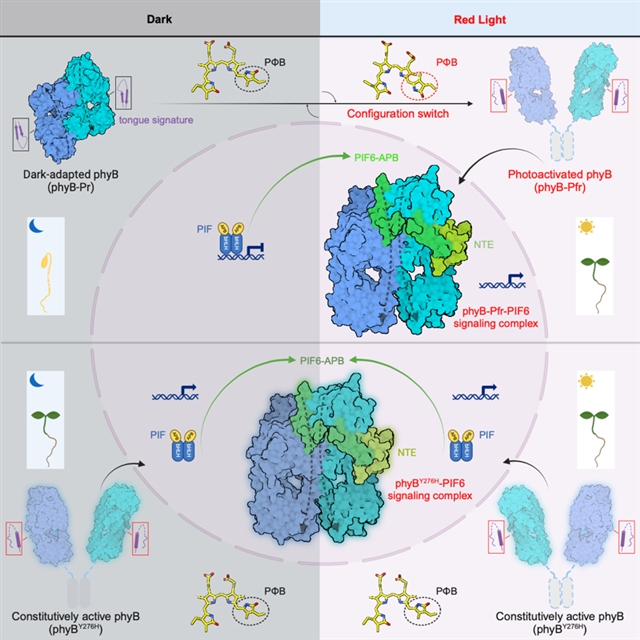
图3. 红光激活phyb并转导pif信号的结构机制模型
研究团队表示,尽管已经揭示了phyb光激活的变构机理以及phyb-pfr特异性识别并结合pifs-apb的机制,还有待于进一步阐释phyb-pfr不对称二聚体的形成机制,并基于此探究系列激活态phyb与下游因子的互作方式,为phyb蛋白在作物性状精准改良以及“光遗传学”工具开发等研究提供更充足的理论支持。(来源:科学网)
相关论文信息:
特别声明:本文转载仅仅是出于传播信息的需要,并不意味着代表本网站观点或证实其内容的真实性;如其他媒体、网站或个人从本网站转载使用,须保留本网站注明的“来源”,并自负米乐app官网下载的版权等法律责任;作者如果不希望被转载或者联系转载稿费等事宜,请与我们接洽。