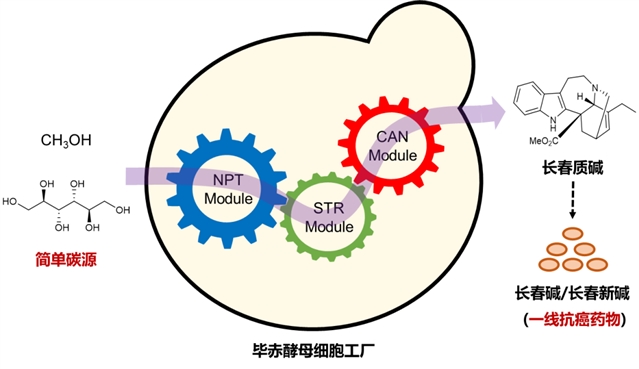
北京时间2023年1月10日,浙江大学连佳长课题组在nature synthesis期刊发表研究论文“biosynthesis of catharanthine in engineered pichia pastoris”,首次报道了利用毕赤酵母细胞工厂来高效合成长春质碱。
继keasling教授团队在nature报道了长春碱前体(长春质碱和文多灵)从头合成的突破性成果之后,浙江大学连佳长课题组在nature synthesis发表研究论文,首次报道了利用毕赤酵母细胞工厂来高效合成长春质碱。长春质碱和文多灵可偶联合成临床应用最为有效的抗癌药物——长春碱。然而,这些植物来源次级代谢产物的天然含量低、结构复杂,阻碍了其大规模的植物提取或化学合成,导致产量低、成本高。本研究以毕赤酵母为底盘细胞重构长春质碱合成途径,实现了从简单碳源到高附加值天然产物的从头合成(图1)。
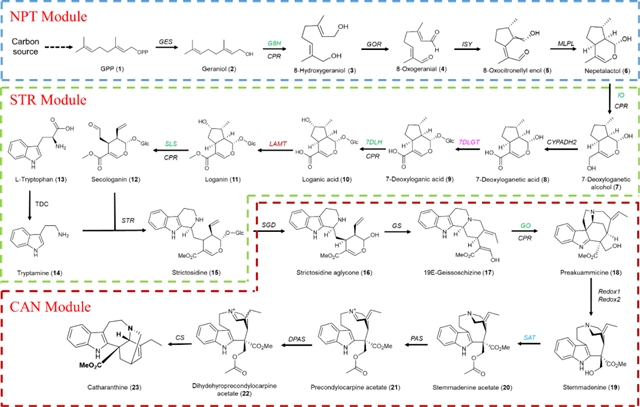
图1:在毕赤酵母中分模块构建及优化长春质碱从头合成途径
▌1. 筛选和表征稳定的整合位点
首先,在毕赤酵母基因组上筛选得到了31个整合位点,以红色荧光蛋白基因为报告基因,测试了各个整合位点的整合效率,并对具有高整合效率的整合位点进行了表达强度测试、稳定性分析和生长分析。31个整合位点中有24个整合位点的整合效率达到了80%以上,且荧光强度和位点存在相关性(图2)。在非选择性培养基中连续转移30代后,所有菌落均呈现红色荧光,表明异源基因表达盒在毕赤酵母基因组的稳定性。此外,对多重基因组整合效率进行了评估,三位点同时整合的效率高于60%。
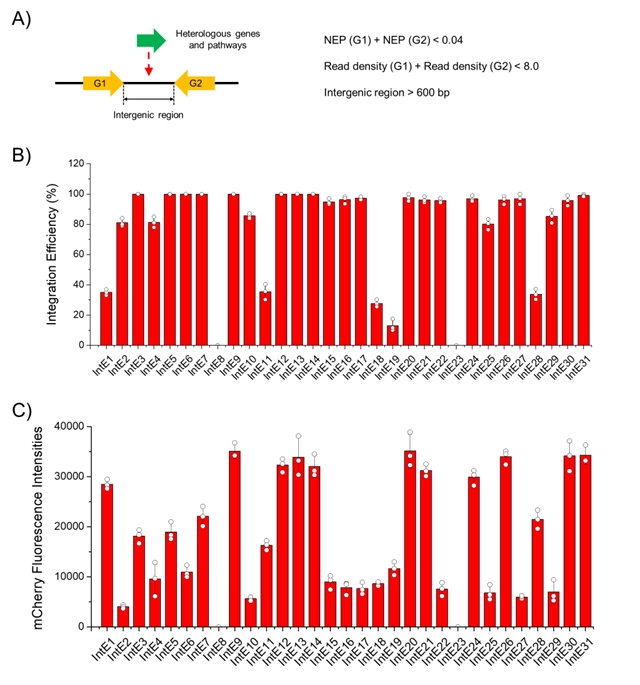
图2:整合位点的挖掘与表征
▌2. 异胡豆苷到长春质碱的生物合成
考虑到长春质碱合成途径的复杂性,将其分成三个功能模块(can模块、str模块和npt模块)进行重构和优化。首先,重构了can模块(can4a),在外源添加异胡豆苷的条件下,长春质碱的产量为125 μg/l(图3a和3b)。crpas是can模块的关键蛋白,将其与mbp融合表达能有效提高长春质碱的产量(mbp3-tpas),达到270 μg/l(图3c)。由于缺乏途径中绝大部分中间体的检测方法,通过增加can模块基因的拷贝数来鉴定途径的限速酶,可以得出sgd、gs、pas、dpas和cs为途径的主要限速酶。在基因组上增加这些限速酶基因的拷贝数,得到菌株can7,当体外添加4.5 mg/l异胡豆苷时,长春质碱的产量可达1.1 mg/l(图3d)。
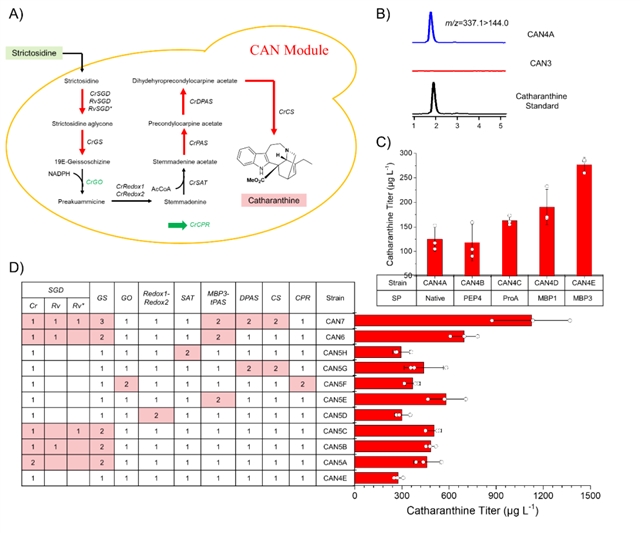
图3:can模块的构建与优化
▌3. 荆芥醇到长春质碱的生物合成
在菌株can7的基础上,引入str模块得到菌株can9(图4a)。在外源添加150 mg/l荆芥醇时,长春质碱的产量为1.2 mg/l。通过增加str模块基因的拷贝数,发现crstr和crsls是主要限速酶,增加其拷贝数(can11),长春质碱的产量为2.7 mg/l(图4b)。
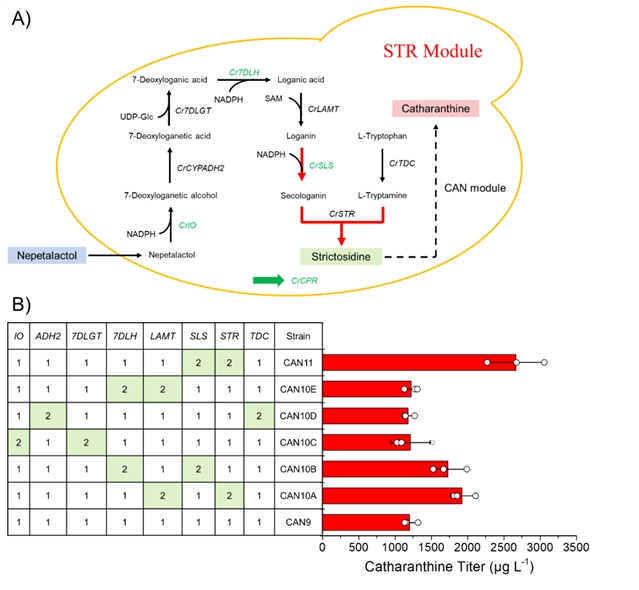
图4:str模块的构建与优化
▌4. 长春质碱的从头生物合成
在菌株can11的基础上,引入npt模块得到菌株can14(图5a),长春质碱的产量仅为1.6 μg/l。在菌株can14中引入mlpl(乳胶蛋白类基因),额外引入由组成型启动子驱动的crg8h和crges-scerg20ww,过表达转录激活因子mxr1,构建得到菌株can17,长春质碱的产量提高到20.3 μg/l(图5b)。当甲醇作为唯一的碳源和能源时,超过70%的碳代谢通量流向了异化途径(生成能量和二氧化碳),而不是同化途径(生物合成),因此尝试引入非抑制性碳源,发现甲醇与海藻糖或甘露醇共培养时,长春质碱的产量可以提高到65 μg/l(图5c)。
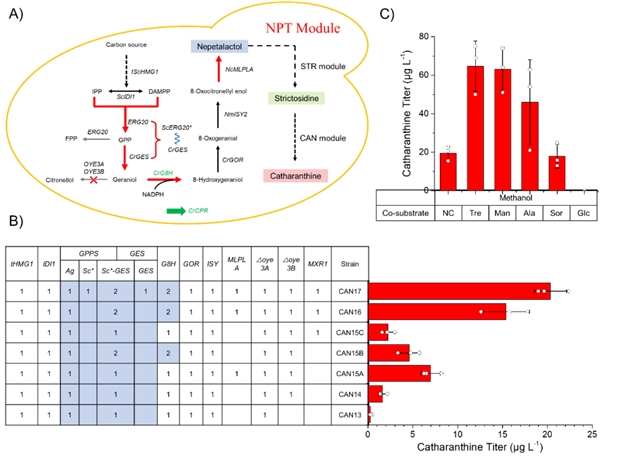
图5:npt模块的构建与优化
▌5. 底盘细胞的代谢工程改造
最后对底盘细胞进行改造。过表达酿酒酵母来源的腺苷甲硫氨酸合酶提高sam供应,可提高长春质碱产量~2倍(can18a)。过表达cyb5提高p450的电子传递效率,使长春质碱的产量提高~1.7倍(can18b)。将这些代谢工程改造策略结合起来构建can19,其长春质碱的产量提高了~6倍(图6a),在1 l发酵罐中进行补料分批发酵,产量可达2.57 mg/l(图6b)。
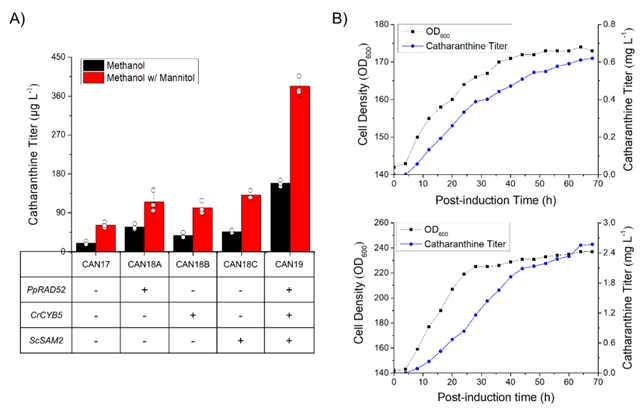
图6:菌株的代谢工程优化与高密度发酵
▌总结
以异胡豆苷和荆芥醇作为节点,将长春质碱合成途径分成三个功能模块,在毕赤酵母内分模块依次进行整合和优化,首次在毕赤酵母实现了长春质碱的从头合成。构建的毕赤酵母工程菌株 can19在摇瓶和发酵罐上的产量分别为0.38 mg/l和2.57 mg/l。长春质碱途径是迄今为止在非模式菌株中构建的最为复杂的生物合成途径,证明了毕赤酵母作为合成植物天然产物细胞工厂的优势和潜力。
本研究得到了国家重点研发计划(2018yfa0901800和2021yfc2103200)、国家自然科学基金(22278361)和浙江省杰出青年基金(lr20b060003)等项目的支持。(来源:科学网)
相关论文信息: